module 18
Important weeds of Australia and southern Asia
Objectives
This module is designed to inform you of some of the important weeds in Australia and southern Asia. At the end of this topic you will be familiar with several of these weeds, their biology and ecology, and their management.
Module Outline
- Parthenium hysterophorus L.
- Lantana camara L.
- Eichhornia crassipes (Mart.) Solms-Laubach
- Jatropha gossypiifolia L.
- Andredera cordifolia L.
- Mikania micrantha H.B.K
Parthenium hysterophorus L.
Name
Parthenium hysterophorus L. (parthenium weed) is a member of the tribe Heliantheae of the Asteraceae, an extremely diverse family with a cosmopolitan distribution. The genus Parthenium contains 15 species which are native to North and South America. Parthenium hysterophorus is the only member of the genus naturalised in Australia.

Figure 18.1 Parthenium hysterophorus
Description
Parthenium weed is an erect and much branched annual, or ephemeral, herb known for its vigorous growth that can overtake other species. It can grow up to 2 m high though most individuals do not exceed 1.5 m.
The flower heads of parthenium weed are composed of many florets formed into small white capitula that are 3-5 mm in diameter. Each head consists of five fertile ray florets and about 40 male disc florets, hence only ray achenes are produced. The first capitulum forms in the terminal leaf axil, while subsequent capitula form progressively down the stem on lateral branches emanating from the axils of lower leaves. Thousands of flower heads, forming in branched clusters at the top of the plant, may be produced on a plant during flowering.
The diaspore is a cypsela with two sterile florets adhering as ‘wings’, and is usually referred to as an achene. These appendages act as air sacs which increase the air-mobility and floatation of the diaspore, therefore aiding the dispersal of the seed. Diaspores vary between 2-3.5 mm in length and are about 2 mm wide. The seed is black, obovate in shape, flattened and crowned by a pappus of orbicular scales.
Intraspecific variation
In North and South America there seem to be two distinct races of parthenium weed, which have been termed the `South American’ and `North American’ races. The `North American’ race has been introduced into Australia, and many other parts of the world. The `South American’ race differs from the `North American’ race as it has larger flower heads and disc florets, yellow petals and less development of axillary branches. Hymenin is the dominant sesquiterpene lactone found in plants of the `South American’ race, whereas parthenin is dominant in the `North American’ race. These differences in the chemistry and morphology of populations of parthenium weed in North and South America indicate the possibility of the existence of several forms, subspecies, or perhaps even species.
Of the two separate introductions of parthenium weed into Australia, plants from the Clermont (central Queensland) introduction seem to be much more aggressive than those from the Toogoolawah (south-east Queensland) introduction. Recently, growth room studies have highlighted differences in the growth and phenology of these two biotypes, and genetic studies have also detected differences between plants from both introductions. In particular, plants from the Clermont introduction grew taller, had a greater biomass and produced larger leaves than plants from the Toogoolawah introduction which were grown in the same conditions.
Distribution
Australia
Parthenium weed occurs in Queensland, New South Wales and the Northern Territory. All of the major infestations of parthenium are found in the sub-coastal regions of central Queensland, in areas with an average annual rainfall of 500-700 mm. Scattered infestations periodically occur throughout the eastern half of central and southern Queensland and into the sub-coastal regions of New South Wales. Parthenium weed is prominent where substantial development of brigalow scrubs has occurred. The areas of land primarily affected are beef producing pastures, but the weed is also spreading into cropping areas and the sugarcane growing areas nearer the coast. The potential distribution of parthenium weed in Australia is much wider than at present, and it has the potential to become a weed of significance throughout the warm and temperate, humid and sub-humid regions of Australia.
Parthenium weed’s distribution seems to be limited to areas that do not experience extremes of temperature (<50oC, >40oC) during the period when there is enough rainfall to permit growth. It has also been suggested that the distribution of this weed may also be limited by heavy shading (>80% shade) and so it should not be a problem in closed forest habitats in Australia.

Figure 18.2 Distribution of Parthenium hysterophorus in Australia (Australia’s Virtual Herbarium 2006)
Outside Australia
Parthenium weed probably originated in the area surrounding the Gulf of Mexico or in central South America. In North America, South America and the Caribbean it is widespread, and has probably spread from its original range as a result of recent anthropogenic disturbances. This spread is most apparent in the Caribbean where it has spread from its previous distribution to all Commonwealth Caribbean countries.
From North America parthenium weed has been introduced to the Republic of South Africa, Ethiopia, Madagascar, Kenya, Mozambique, Mauritius, Rodriguez, the Seychelles, Israel, India, Bangladesh, Nepal, China, Vietnam, Taiwan, Australia, and many Pacific Islands. It has had by far the greatest impact in India where it is widespread.
Habitat
Parthenium weed is especially prolific in disturbed situations and occurs in naturally disturbed areas, as well as those that receive constant traffic from vehicles or livestock. It shows a marked preference for black, alkaline, cracking, clay soils of high fertility, however it will grow on a wide variety of soils. On other soil types it usually requires a more severe soil disturbance to establish, and rarely forms the extensive stands found on the heavy black soils.
In Australia the weed predominantly inhabits pasture areas and has had little impact in cultivated areas in contrast to the situation in the Americas and India. Parthenium weed is vigorous in both established and developing pastures. However it will not establish where there is a high level of ground cover. The initial occurrence of parthenium weed in a new area is frequently along roadsides, and it is from this foothold that it spreads laterally and extensively into agricultural land.
Growth and development
Morphology
Parthenium weed, in its early stages of growth, exists as a rosette and so requires a suitably open area in which to establish. This rosette spreads radially very close to the ground and interferes with the emergence of other seedlings. The stem of the weed then elongates rapidly and starts branching at the apex. Mature plants are much-branched, and axillary branches also form down the stem as the plant gets older. This growth form coupled with the weed’s high growth rate, allows it to be very competitive and enables it to exclude the growth of other species.
Phenology
Parthenium weed is able to germinate, grow and flower over a wide range of temperatures and photoperiods, and can complete its life cycle at any time of the year in warmer climates. The main season of growth, however, is during the summer months (November – March) when rain is usually more abundant. Four or more successive cohorts of seedlings may emerge at the same site during a good growing season.
Reproduction
Floral biology
Flowering can be initiated as early as 4 weeks after seedling emergence. In developmental studies conducted on North American plants, the time from the initial appearance of the first flower bud to the production of a mature inflorescence and dispersal of the first achenes was found to be about 30 days, while the time from pollination to achene maturation is only about 14 days. An average of 150,000 – 350,000 pollen grains are produced in each flower head, and as thousands of flower
heads can be present on each plant, pollen production by each plant is extremely large; i.e. 15 – 850 million pollen. In the USA and India airborne pollen from parthenium weed has been detected in significant amounts at a variety of altitudes (2–915 m) and distances (up to several km) from populations of the weed.
Seed production and dispersal
Parthenium weed is a very prolific seed producer and will continue to flower and fruit profusely until senescence. The achenes are shed gradually or retained on the inflorescence until after senescence. Each flower head produces 5 achenes of uniform size, weighing from 0.4 to 0.8 mg.
A typical mature plant can produce over 15,000 achenes. In India an average of 15 parthenium weed plants per m2 were present in a stand of the weed and in excess of 300,000 achenes per square metre could be produced under these circumstances. Soil seed bank studies of parthenium weed in Australia have detected between 1,500 and 34,000 per m2 in pastures. In these studies parthenium weed seeds represented between 47 and 87% of the entire soil seed bank
Dispersal of parthenium weed occurs locally by wind and water. Wind transport is usually only in the order of a few metres, but whirlwinds can carry large numbers of the light seed for considerable distances. Dispersal of achenes by water is important as indicated by large populations of the weed spreading along waterways in central Queensland. Native animals, livestock and feral animals are also involved in the dispersal of parthenium weed seed.
Parthenium weed achenes are capable of being transported long distances in mud and debris. In the majority of cases of long distance dispersal, seeds are transported on motor vehicles or machinery, on livestock, with crop and pasture seed, or in fodder. As a result, new infestations of the weed may appear thousands of kilometres from the nearest plants.
Physiology of seeds and germination
It has often been assumed that parthenium weed seeds possess no primary dormancy mechanism. However, research has demonstrated that water soluble germination inhibitors (i.e. parthenin and phenolic acids) are present in the accessory structures and seed coat of parthenium weed seeds, and these inhibitors need to be leached before maximum germination is reached.
As is the case with achene dormancy, little is known about the longevity of parthenium weed achenes. In one study it was found that, of the seeds recovered, germination declined from 66% after one week of burial to 29% after burial for two years. Depth of burial did not affect subsequent germination percentage. These germination tests were conducted in the dark, and at constant incubation temperature,
so adequate conditions may not have been present for breaking any induced dormancy that may have been present. It is possible, therefore, that the viability of these seeds may have been much higher than was reported since they may have been dormant rather than non-viable. Some field evidence exists that parthenium weed achenes can remain viable after being buried for at least 4-6 years.
The longevity of surface-lying achenes seems to be rather short. Research shows that most unburied parthenium weed achenes germinate, become non-viable or are harvested by ants, within two years.
Population dynamics
In India, seedlings of parthenium weed were recruited in three successive cohorts after the first monsoon rains. Seedling density and survivorship to maturity declined in successive cohorts and the pattern of recruitment and population density was remarkably similar in two successive years. The average recruitment for the two years was 110 plants per m2 of which 14 plants per m2 were able to reach maturity. The results indicated that the established cohorts adversely affected the growth, and probably survivorship, of the latter accessions through resource competition.
Importance
Parthenium weed is an extremely prolific weed that has the ability to spread quickly and colonize areas. In the pastoral regions of central Queensland parthenium weed is the dominant species under certain conditions. It excludes all beneficial forage plants in a monoculture of non-nutritious, vegetable matter in which it is impossible to sustain cattle. This causes a catastrophic drop in the productivity of pastures, in which the carrying capacity can be reduced to negligible proportions.
The worst infestations in Australia occur in areas that have undergone clearing or have been overgrazed. Under continued heavy grazing, parthenium weed will come to dominate native pastures on many soil types in central Queensland. The presence of stock on newly cleared country is the most consistent pair of factors that heavily infested areas have in common. By 1991 parthenium weed was present throughout 170,000 km2 of Queensland’s prime grazing country, or 10% of the entire state. Annual losses to beef producers as a result of parthenium weed were in the vicinity of $A16.5 million. These losses were due to reduced stock numbers and liveweight gains, and additional production and control costs.
In India parthenium weed has encroached on cultivated fields and grasslands, and can reduce the grass forage to 10% of its normal yield. Parthenium weed is also a serious weed of crops and orchards throughout India as well as in many other countries of the world.
In Israel parthenium weed has been encountered as a weed in tomato (Lycopersicon esculentum Miller), cotton (Gossypium spp.) and forage fields, while in the Caribbean parthenium weed is regarded as the fourth most serious weed species and was found to be one of the main weed species in a selection of 18 different crops grown throughout Jamaica (Hammerton 1974). In North America the weed has been recorded in a range of crops including sugar cane (Saccharum officinarum L.), maize (Zea mays L.), cotton, sorghum (Sorghum bicolour (L.) Moench), onions (Allium cepa L.), and citrus orchards, although not in sufficient numbers to significantly affect production.
Parthenium weed seed is also a contaminant of produce (i.e. grain, pasture seed and forage), the sale and movement of which can be subsequently restricted. The weed has spread from Queensland to New South Wales as a result of contaminated pasture seed. Parthenium weed achenes are also spread by harvesters and other agricultural or industrial machinery. This has led to the construction and use of wash-down facilities, which are both costly and an inconvenience to those who are forced to use them.
Parthenium weed is also an environmental weed which can cause a total habitat change in native grasslands, the understorey of open woodlands and along rivers and floodplains. In areas that are regularly flooded, parthenium weed is difficult to control because the grass cover is killed as a result of submersion and the weed then has no competition.
One of the major detrimental effects of parthenium weed, and a reason for its aggressiveness, is its allelopathic effect on other plants. Allelopathy is the direct or indirect injurious effect of one plant upon another through the exudation of phytotoxic chemicals.
Parthenium weed is usually avoided by stock - it is toxic to animals. However, in situations where the weed forms almost pure stands animals may consume sufficient quantities of it. Studies in India on the toxicity of the weed in cattle and buffaloes have shown that a significant amount (10-50%) of parthenium weed in the diet can kill these animals within 30 days. In such cases the cattle often developed dermatitis with pronounced skin lesions, became highly emaciated, and eventually died due to the rupture of tissues causing haemorrhages in their internal organs.
One of the most detrimental effects of parthenium weed is the human health hazard that it poses. Those who have continued close contact with the weed can develop allergic eczematous contact dermatitis. Parthenin is the causative agent of this reaction, and is one of the very reactive toxic classes of compounds known as sesquiterpene lactones. There has been an epidemic of hundreds of cases of parthenium weed dermatitis in India and several cases have been reported from the U.S.A. The contact allergy can be developed from repeated contact with the weed or its disseminated parts, and can be perpetuated in sensitized individuals by airborne pieces of dried plant material, such as trichomes. Patients with severe dermatitis suffer fatigue and weight loss, and about 12 deaths have occurred in severely affected patients.
In Queensland several sensitized individuals have had to change residences and leave employment as a result of dermatitis caused by the plant. Data collected from a survey conducted in Queensland indicated that 10% of workers on properties in the parthenium weed infested area had developed visible skin allergies to parthenium weed.
The pollen of parthenium weed has also been observed to cause allergic rhinitis (hayfever) and allergic bronchitis (asthma) in humans. In research conducted in the USA, parthenium weed pollen is a cause of allergic disease whose importance has largely been overlooked.
Weed management
Herbicides
Herbicides are available that provide effective control of parthenium weed in almost any situation. Unfortunately, after the successful application of herbicides parthenium weed will usually reappear from seed in the soil. Residual herbicides help to overcome this problem to a certain extent. However, the spraying of plants before they set seed is critical to obtain long term control and a close watch should be kept on treated areas for at least two years. Such control is only feasible in cultivation or in small areas of pasture. Where parthenium weed covers very large areas chemical control is unlikely to be economically viable. In general it is better to spray with a mixture of atrazine and 2,4-D, as 2,4-D kills existing plants, while atrazine provides long term residual activity, but has very little knockdown effect by itself. Dense infestations will often require herbicide treatment in conjunction with pasture management to attain effective long-term control.
Other treatments
Pasture and grazing management
As there seems to be a definite relationship between the invasion of parthenium weed and the vigour of pastures, it is apparent that property management is fundamental to the control of this weed. Pastures should not be grazed heavily as this increases the likelihood of invasion by parthenium weed, and the severity of existing infestations. Rehabilitation of infested pastures requires them to be spelled for at least one full summer, and they must also be grazed very lightly in the following winter. Pasture species may be sown to encourage the restoration of the pasture, but newly sown areas must also be spelled to allow the pasture to establish. Once cover is established, stocking rates have to be carefully adjusted according to season and rainfall to maintain grass dominance. Badly infested areas can be fenced off and destocked to prevent seed spreading to parthenium-free areas.
Physical methods
Fire will produce beneficial results only after the first heavy rains, when the majority of parthenium weed seed has germinated. Burning usually only gives short term control but may be effective when used in conjunction with the sowing of pasture seed and implementation of responsible management practices. However it is often hard to obtain sufficient fuel for a fire once parthenium weed has invaded.
Mechanical treatments such as grading, slashing and ploughing are not considered to be efficient as such methods may aid the spread of parthenium weed achenes. Mowing or slashing also results in the rapid regeneration of plants from lateral shoots close to the ground.
Hygiene
Proper cleaning of cultivating and harvesting vehicles, sowing of uncontaminated seed, and short term quarantine of stock that have been in parthenium weed infested areas will reduce the risk of spreading parthenium weed. Machinery and vehicles that have been in infested areas must be thoroughly cleaned by washing them with a high pressure hose. Stock brought from infested areas should be kept in a holding yard for a few days before transferring them to clean pasture or transporting them elsewhere.
Biological control
Biological control seems to offer the best long-term solution to the parthenium weed problem. However, to date the biological control campaign against parthenium weed in Australia and India has resulted in only limited and inadequate control.
Many species of insects have been found feeding on parthenium weed in its countries of origin (i.e. Mexico, Brazil and Argentina), although most of these are only occasional feeders and have little adverse effect on the plant. Of the species found to be damaging and suitable for introduction into Australia, eight have been released in Queensland since 1980.
The species that are successfully established in Queensland are: the leaf-feeding beetle Zygogramma bicolorata, the stem boring weevil Listronotus setosipennis, the leaf-mining moth Bucculatrix parthenica, the seed-feeding weevil Smicronyx lutulentis and the stem-galling moth Epiblema strenuana. Z. bicolorata was initially very promising as a biocontrol agent for parthenium weed as it breeds rapidly and when present in large numbers can severely defoliate the weed, thereby preventing seed production. However, for many years the population and spread of Z. bicolorata was very low. The beetle is now becoming adapted to central Queensland conditions and is appearing in greater numbers, and having a more significant impact, each year. Z. bicolorata has also been released in India and is starting to have some impact on the weed in the Bangalore area.
Epiblema strenuana has had the most significant impact on parthenium weed in Australia. Parthenium weed has become less competitive and easier to manage than it was prior to this insect’s release. The moth’s larvae form galls in the stems and growing points of parthenium weed and can considerably stunt growth, ultimately reducing seed production. Research has shown that if Epiblema larvae are released onto parthenium weed when it is less than 20 cm in height, in the presence of grass competition, they significantly reduce growth and seed production (T. Priest personal communication).Stands of parthenium weed 1.5-2 m tall were common in central Queensland before E. strenuana was released; now they rarely exceed 1 m in height (McFadyen 1989).
Parthenium weed is a host to two species of pathogenic rust fungi Puccinia abrupta var. partheniicola and P.melampodii Diet. & Holw. in Mexico (Parker 1989). P. abrupta var. partheniicola can be quite damaging, infection with the rust often resulting in a 90% reduction in flower production (Parker et al. 1994). This species was deemed sufficiently host-specific for introduction into Australia and released in 1992, although it is winter acting and the area where parthenium is currently a problem has a summer-dominant rainfall. More recently, Puccinia melanpodii has been released.
Table 18.1: Species released as biological control agents of Parthenium hysterophorus and their establishment in Australia
| Species | Country of Origin | First released | Establishment |
| Insects | |||
| Conotrachelus sp. | Argentina | 1993 | No |
| Listronotus sitosipennis | Brazil/Argentina | 1983 | Yes |
| Smicronyx lutulentus | Mexico | 1980 | No |
| Zygogramma bicolorata | Mexico | 1980 | Yes |
| Stobaera concinna | Mexico | 1983 | Yes - local |
| Buccalatrix parthenica | Mexico | 1984 | Yes |
| Epiblema strenuana | Mexico | 1982 | Yes |
| Platphalonidia mystica | Argentina | 1992 | No |
| Pathogens | |||
| Puccinia abrupta var. | |||
| partheniicola | Mexico | 1992 | Yes |
| Puccinia melanpodii | Mexico | 1992 | Yes |
|
References
Auld, BA, Hosking, J and McFadyen, RE 1983, ‘Analysis of the spread of tiger pear and parthenium weed in Australia, Australian Weeds vol. 2, pp. 56-60.Australia’s Virtual Herbarium, http://www.anbg.gov.au/avh.html. Commonwealth State and Territory Governments of Australia and the Council of Heads of Australian Herbaria, accessed July 25th 2006.
Butler, JE 1984, ‘Longevity of Parthenium hysterophorus L. seed in the soil’, Australian Weeds vol. 3, p. 6.
Chippendale, JF and Panetta, FD 1994, ‘The cost of parthenium weed to the Queensland cattle industry’, Plant Protection Quarterly vol. 9, pp. 73-6.
Dale, IJ 1981, ‘Parthenium weed in the Americas’, Australian Weeds vol. 1, pp. 8-14.
McFadyen, RE 1992, ‘Biological control against parthenium weed in Australia’,Crop Protection, vol. 11, pp. 400-7.
Navie, SC, McFadyen, RE, Panetta, FD and Adkins, SW 1998, ‘Parthenium hysterophorus L’, in Panetta, FD, Groves, RH and Shepherd, RCH (eds.) The biology of Australian weeds, Volume 2. RG and FJ Richardson, Meredith, Victoria, pp. 157-176.
Pandey, HN and Dubey, SK (1988, ‘Achene germination of Parthenium hysterophorus L.: effects of light, temperature, provenance and achene size’, Weed Research, vol. 28, pp. 185-90.
Parsons, WT and Cuthbertson, EG 1992, Noxious Weeds of Australia, Inkata Press, Melbourne, pp. 292-6.
Williams, JD and Groves, RH 1980 ‘The influence of temperature and photoperiod on growth and development of Parthenium hysterophorus L.’, Weed Research, vol. 20, pp. 387-92.
Lantana camara L.
(This description draws heavily on information provided in Swarbrick et al. 1998.)
Name
Lantana camara L. (lantana) was known by at least five polynomial descriptive names, starting with Lantana, Vibernum and Periclymenum, before Linnaeus gave it its binomial name in 1753. He retained Lantana (origin obscure) and described a number of species including camara (a West Indian name) and aculeata (prickly), a species now included within the L. camara complex (L. camara sensu lato). Linnaeus’ L. camara was non-prickly whilst his L. aculeata was prickly; both were described from European garden material (Schauer 1851).

Figure 18.3 Lantana camara L.
Description
Lantana is a brittle, much branched, thicket-forming shrub, normally 2-4 m tall but capable of becoming a liane up to 15 m tall if given support.
The arching, scrambling or prostrate stems are initially 4-angled but become cylindrical and up to 15 cm thick with age. Young stems are hairy and in the weedy forms carry sharp recurved prickles along the angles, whilst those of the non-weedy forms are rounder, more slender and without prickles.
Inflorescences develop in the axils of the leaves. They are compact, flat to dome-shaped and 2-3 cm across and each is supported by a finely hairy 2-8 cm long peduncle with a club-shaped tip. Each inflorescence contains 20-40 sessile flowers, each subtended by a stiff deciduous bract.
Intraspecific variation
Lantana is an aggregate species, derived through horticultural and natural hybridization, selection and somatic mutation from a number of similar and probably closely related (though spatially separated) tropical American species. This has resulted in many hundreds of forms, which are also referred to as cultivars and varieties (Howard 1969).
At least 29 forms have become naturalised in eastern Australia and probably others in the Northern Territory and Western Australia. Most of the weedy forms of lantana in South Africa and India and probably Australia appear to be tetraploids, whilst most of the non-weedy ornamental forms appear to be triploids.
History
Lantana was probably first introduced into Europe about 1640 from Brazil (Howard 1969), where both prickly and non-prickly taxa are considered to be native, common and widespread (Schauer 1851). Introductions continued throughout the 17th, 18th and 19th centuries (Swarbrick 1985) resulting in an aggregate species of mixed parentage (Howard 1969). Lantana became a favourite hothouse plant in Europe in the late 19th century, with 397 new varietal names being added to nurserymen’s catalogues between 1850 and 1900 (Howard 1969). Many of these forms would be distributed to the colonies, including Australia.
The first Australian record of lantana was in 1841 at the old Botanic Gardens in Adelaide, and was followed by many others. The plant was quickly spread northwards along the coast, with the first Australian collections of apparently naturalised lantanas from the Brisbane River in 1861 and near Sydney in 1862 (Swarbrick 1985).
Distribution
Australia
Lantana is widely distributed throughout coastal and sub-coastal areas from Ulladulla in New South Wales to Cape Melville in Queensland, with very scattered infestations in the Northern Territory, Western and South Australia (Figure 2.4). It is also present as a weed on both Lord Howe and Norfolk Islands (Swarbrick 1985).

Figure 18.4 The Australian distribution of lantana (Australia’s Virtual Herbarium 2006)
Outside Australia
Lantana is widely grown as an ornamental shrub throughout the tropics, subtropics and warm temperate zones. It is established as a weed throughout the tropics and subtropics, from southern USA and the Mediterranean in the north to South Africa and the northern tip of New Zealand in the south.
Habitat
Lantana grows well over a wide range of warm temperate, subtropical and tropical climates. It does not grow at temperatures below 50C and the shoots are sensitive to frost.
In south-eastern Queensland it is similarly found in moist coastal areas that naturally support rainforest, and drier eastern slopes of the Dividing Range which support open eucalypt woodland, but is not found west of the Diving Range where both drought and frost occur.
Lantana grows best on rich organic soils developed under rainforests, although it is also reported to tolerate poor soils and almost pure sands. In the wet tropics of northern Queensland lantana is mostly confined to the deeper, moister and better drained metamorphic and granitic soils, not occurring where soils are shallow and severely drained.
Growth and development
Morphology
Lantana is very plastic in its responses to light intensity and support. Seedling densities vary from less than one to several per square metre. Over a number of years some plants dominate and shade out the rest, with each dominant plant eventually occupying several square metres.
New shoots constantly arise from the base of large healthy plants, whilst older shaded stems lose vigour and die. Seedlings germinating under such clumps usually fail to establish.
The presence of a low-branched tree or other support allows lantana to scramble upwards to a height of 10-15 m, in which case the lower stems lose their branches and assume a flexible liane-like form.
Phenology
Parsons and Cuthbertson (1992) reported that lantana seedlings germinate throughout the year provided sufficient moisture is present, with a peak after the first summer rains. Initial growth is slow, until the root system is established, after which it increases as the season advances. They reported that the first flowers are produced in the field during the second summer, although flowers can be produced six months after germination in the greenhouse.
Reproduction
Floral biology
Parsons and Cuthbertson (1992) reported that under good conditions lantana plants flower from early summer until March or April of the year after germination and every summer thereafter from September or October until March or April.
Seed production and dispersal
Since there are normally two inflorescences per node during active growth, the total number of fruits produced by a lantana plant under good growing conditions is several thousand per square metre per year.
Lantana seeds may either be transported away from the plant or fall beneath the parent plant. Most fruits are thought to be dispersed by birds, often locally but also to distances of up to 1 km or more. The seeds resist digestion during passage through the vector, and initial infestations from bird-dispersed seeds are most commonly seen under fences, trees and other obvious perches.
Physiology of seeds and germination
Whilst investigating the effects of the lantana seed fly Ophiomyia lantanae (Froggatt) on the germination of lantana seeds from Natal, Graaff (1986) recorded a very low germination rate even in uninfested fruit. Of 1200 seeds examined only 48% had fully formed embryos. Breaking the dormancy (possibly caused by the woody seed coat) by
180 days of dry storage increased germination from 29 to 47% after 85 days. The lantana seed fly also affected the germination percentage of infested fruits (see response to biological control below).
Importance
Pastures
The main economic damage caused by lantana is through decreased productivity of pasture land. Lantana readily invades uncultivated pastures, and unless controlled spreads to exclude useful native grasses and fodders. The plant is unpalatable and scarcely eaten, and the trampling and deposition of dung by large hoofed animals is instrumental in the germination and establishment of new seedlings.
Lantana contains a number of chemicals toxic to animals, especially the triterpene lantadenes A and B. Whilst ungulates are primarily affected, there have been some cases of human poisoning. All forms of lantana are thought to be toxic, although the toxicity varies considerably between forms.
Nearly 4 million hectares of pasture are thought to be infested with lantana in Australia (Parsons and Cuthbertson 1992), mostly in coastal and subcoastal Queensland and New South Wales.
Annual losses due to lantana in pastures are estimated to be $A7.7 million, comprising 1500 cattle deaths ($A0.5 million), 4.5% reduced performance ($A2.0 million), 7.3% loss of pasture ($A3.0 million) and $A2.2 million in control costs (Culvenor 1985).
Natural ecosystems
Lantana is a serious invader of disturbed ecosystems conserved for their natural and semi-natural attributes throughout its Australian and overseas distribution. It is among the most widespread and obvious of the exotic species in the Wet Tropics Heritage Area of northern Queensland where it invades disturbed rainforest.
Lantana is also an aggressive invader of natural ecosystems in many other countries, for example, the Galapagos Islands, many Pacific islands and South Africa.
Forestry
Lantana is a serious pest of commercial hoop pine plantations in coastal south-eastern Queensland, competing with tree seedlings for light, interfering with access throughout the life of the plantation, and increasing both management and harvesting costs and fire hazard.
Weed management
Herbicides
Many chemical control trials have been conducted against the various forms of lantana in Australia, South Africa, Zimbabwe, India, Taiwan and the USA. The comparative efficacy of various herbicides and factors affecting their efficacy are discussed elsewhere (Swarbrick et al. 1998).
Other treatments
Although the use of fire to control lantana in pasture is widely recommended (Goodchild 1951, Saint-Smith 1964, Bartholomew and Armstrong 1978), very little experimental work has been published on its use and effects. Fire is generally insufficient by itself for lantana control, and additional treatments including sowing competitive pasture species, controlled grazing and follow-up burns and chemical treatments are usually required. Little information is available on the seasonality of burning, the effect of meteorological conditions at the time of the burn or the occurrence of rain after burning, the quality and type of fuel, whether a ground or crown fire is preferable, or time of reintroduction of stock after the fire.
Several authors have reported the effects of mechanical clearance of lantana in pastures (Goodchild 1951, Saint-Smith 1964, Bartholomew and Armstrong 1978). All have shown that cultivation gives very effective control of lantana but that effective pasture establishment and management are essential and that some follow-up control is generally required.
Biological control
Surveys of the phytophagous insect fauna of L. camara and closely related species have yielded 544 species in North and Central America and 345 species in Brazil. Most are probably polyphagous and thus unsuitable as biological control agents.
Lantana was the first weed against which biological control was attempted. Koebele in 1902 collected 23 insect species in Mexico and shipped them to Hawaii, where eight species were eventually established. Julien (1992) lists 32 insect species that have been deliberately released in 25 countries. Since then the leaf-feeding chrysomelid beetle Charidotis pygmaea and oecophorid moth Ectaga garcia have been released in Queensland and the leaf-mining chrysomelid beetles Uroplata girardi and Octotoma scabripennis and leaf sap-sucking bug Teleonemia scrupulosa have been released in the Solomon Islands.
Despite this massive effort lantana remains one of the world’s worst weeds and thus its biological control can be considered to have been unsuccessful, even though levels of control ranging from minor to significant have been achieved in some countries.
Ophiomyia lantanae was one of the original insects found on lantana by Koebele and is now established in nine of the eleven countries in which it has been released. It oviposits in green fruits, where larvae feed on the fleshy pulp and may enter the seed and feed on the embryo.
The tingid bug T. scrupulosa is established in twenty countries. It oviposits in stems, petioles and peduncles, most often where adults have flowers on which to feed.
References and further reading
Australia’s Virtual Herbarium, <http://www.anbg.gov.au/avh.html>, Commonwealth State and Territory Governments of Australia and the Council of Heads of Australian Herbaria, accessed July 25th 2006.
Bartholomew, BL and Armstrong, TR 1978, ‘A new look at lantana control’, Queensland Agricultural Journal, vol. 104, pp. 339-44.
Culvenor, CCJ 1985, ‘Economic loss due to poisonous plants in Australia’, in Plant Toxicology: Proceedings of the Australia – USA Poisonous Plants Symposium, Queensland Department of Primary Industries, Brisbane.
Goodchild, NE 1951, ‘Control of lantana by cultural methods in the Mackay district’, Queensland Agricultural Journal, vol. 72, pp. 11-16.
Graaff, JL 1986, ‘Lantana camara, the plant and some methods for its control’,South African Forestry Journal, vol. 136, pp. 26-30.
Howard, RA 1969, ‘A checklist of cultivar names used in the genus Lantana’, Arnoldia, vol. 29, pp. 73-109.
Julien, MH (ed.) 1992, Biological Control of Weeds: A World Catalogue of Agents and their Target Weeds, CAB International Institute of Biological Control, UK.
Parsons, WT and Cuthbertson, EG 1992, Noxious Weeds of Australia, Inkata Press, Melbourne.
Saint-Smith, JH 1964, ‘Pasture weeds of the wet tropics’, Queensland Agricultural Journal, vol. 90, pp. 289-300.
Schauer, JC 1851, ‘Verbenaceae’, in Martius, CFP (ed.) Flora Brasiliensis Volume 9, R Oldenberg, Leipzig.
Swarbrick, JT 1985, ‘History of the lantanas in Australia and origins of the weedy biotypes’, Plant Protection Quarterly, vol. 1, pp. 115-21.
Swarbrick, JT, Willson, BW and Hannan-Jones, MA 1998, ‘Lantana camara L.’, in Panetta, FD, Groves, RH and Shepherd, RCH (eds.) The Biology of Australian Weeds, Volume 2. RG and FJ Richardson, Meredith, Victoria, pp. 119-140.
Eichhornia crassipes(Mart.) Solms-Laubach
This description draws heavily on Wright and Purcell (1995).
Name
Eichhornia crassipes (Mart.) Solms-Laubach (Pontederiaceae) Synonym: E. speciosa Kunth. Common name: water hyacinth

Figure 18.5 Eichhornia crassipes
Eichhornia crassipes is an erect floating perennial herb, reproducing from stolons and by seed. Stems are either erect (inflorescence stalk), to 60 cm long and bearing flowers or horizontal (stolons), about 60 cm long and producing new plants from terminal buds; glabrous.
There are two leaf forms ranging from plants having swollen or bulbous petioles up to 25 cm long but usually less, to plants having slender or non-bulbous petioles to 60 cm or longer. Petioles contain variable amounts of air enabling the plant to float on water. Leaf blades or laminae are smooth, entire, glossy and with semi-parallel veins.
Flowers are bluish purple, funnel-shaped, 4-7 cm diameter, with six lobes or petals, the upper lobe with a yellow blotch in the centre surrounded by darker purple; produced in spikes about 15 cm long on stalks about the same length, each spike consisting of about eight (range 3-35) flowers arranged spirally and subtended by two leafy bracts; individual flowers last only a few days.
The root is fibrous, feather-like with many laterals, dark-coloured, emerging from a rhizome or crown, trailing freely in water to a depth of 1 m or more.
E. crassipes is usually free-floating. Other members of this aquatic family are always firmly rooted in the substrate.
Intraspecific variation
The varying growth form of Eichhornia crassipes ranges continuously from plants having bulbous, ‘float’ petioles to plants with petioles that are slender and non-bulbous. Plants with non-bulbous petioles are commonly found in dense infestations where, presumably, space is limited, intraspecific competition is high and neighbouring plants support each other. Plants with bulbous petioles occur at the open water margins of dense infestations or in infestations of low plant density.
History
Water hyacinth is presumed native to Brazil and other central and south American countries (Penfound and Earle 1948) although studies of its floral polymorphism suggest a more restricted Amazonian origin (Barrett and Forno 1982).
Distribution
It was first recorded in Australia about 1894 from localities in Queensland and New South Wales. It is probable that it was introduced as an ornamental and later discarded into creeks and rivers. By 1900 it was firmly established.
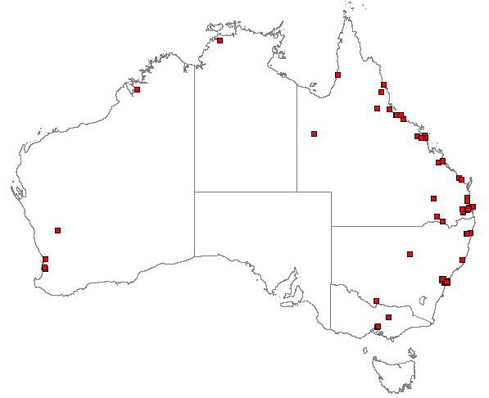
Figure 18.6 Distribution of E. crassipes in Australia (Australia’s Virtual Herbarium 2006)
Habitat
Eichhornia crassipes is a potential threat to all freshwater bodies in mainland Australia. It can sometimes survive rigorous winter conditions in Japan (Ueki and Oki 1979) and the USA (Penfound and Earle 1948), and could possibly survive as far south as Tasmania. Frost causes die-back of the leaves and the plant may die if the upper part of the rhizome is frosted, however seeds can successfully survive the winter.
Eichhornia crassipes can withstand a wide range of changes to the substrate which is usually still or slowly moving fresh water. Plants will grow in mud and can survive for several months on a substrate of low moisture content.
Frequently E. crassipes is the dominant plant species in a fresh water community and may be the only floating species present. As an invader of aquatic communities, it reduces or eliminates other plant species, both native and introduced. In its native range, E .crassipes infests coastal lowlands and the margins of lagoons and slow-moving waters. Seldom are water hyacinth mats as dense as those of infestations in Australia, and then only where there have been hydrological disturbances by man.
Growth and development
E. crassipes is a perennial. During autumn and winter the plant grows slowly and if frosted, leaves die back to the junction of the petiole and the rhizome. The rhizome is thought to be the main storage area for starch (Penfound and Earle 1948). These authors suggested that after it reaches a certain length, decay is equal to growth and the rhizome maintains an almost constant length.
In Australia, plants flower mainly in spring and summer and seed can be set after any period of flowering. Perennation also occurs as a result of extended dormancy of seeds. Seedlings commonly develop on wet mud as plants with bulbous petioles. If seedlings and/or mature plants rooted in mud are submerged by rising water levels, they can absciss from the roots and float to the water surface where they rapidly produce a new root system.
Reproduction
Floral biology
Although the family Pontederiaceae is trystilic, populations of E. crassipes are usually dominated by one style form, and trystily is generally non-functional (Barrett 1977). All three style forms are present in the Amazon Basin and the Mato Grosso of Brazil (Barrett and Forno 1982) but only two occur in Australia: one with the style longer than all the anthers and the other with the style intermediate in length between the two sets of anthers. Populations may contain one or both flower forms. Mid-stylic flowers are the most common.
Seed production
Both long- and mid-stylic flowers set seed in Australia. After flowering, the flower stalk bends until the flower cluster is completely inverted and if unobstructed, becomes submerged. Seeds can mature above or below the water surface and each seed capsule may contain up to 300 seeds (Manson and Manson 1958). Seeds sink following release from the capsule. There are no obvious mechanisms for dispersal.
Physiology of seeds and germination
Seeds are often the source of a new infestation or a reinfestation (Pieterse 1978). Seed has been reported to remain viable for 5 – 20 years (Manson and Manson 1958, Das 1969, Matthews et al. 1977). There are several conflicting reports on the requirements for seed germination. Experimental studies by Obeid and Tag el Seed (1976) showed water depth, soil type, bank deposits and light and/or temperature were important and they concluded that the germination of seed in its natural habitat would depend on the conditions prevailing at the time.
Germination in the field is usually observed on shorelines of lagoons with fluctuating water levels. Seedlings have also been observed on rotting mats of water hyacinth either on shorelines or floating on lagoons. Seeds may also germinate in very shallow water.
Vegetative reproduction
This is the most common mode of reproduction and is largely responsible for the rapid increase in field populations following colonisation by water hyacinth. Propagation occurs by means of stolons which produce daughter plants at the apices. These new plants quickly produce root systems and on decay or breakage of the connecting stolon, they become independent of the mother plant. A single plant can produce a new infestation.
Importance
Eichhornia crassipes poses problems to water use (domestic and commercial) in many natural watercourses, natural and man-made lakes, irrigation and flood mitigation channels and dams. It can impede water transport, prevent water sports and change the physical and chemical composition of the water resulting in changes to the natural biotic community.
In Australia, water hyacinth usually forms a dense impenetrable cover which alters the micro-environment below the water surface. Many direct and indirect effects on the freshwater environment by water hyacinth mats have been reported, including reductions in pH, the penetration of light and levels of dissolved oxygen and by increases in carbon dioxide levels, resulting in negative effects on communities of aquatic vertebrates, invertebrates and plants (Ultsch 1973).
Weed management
Herbicides
Eichhornia crassipes is very susceptible to diquat and the amine and acid formulations of 2,4-D. These have proved very successful for the control of this plant in single-purpose water systems such as irrigation canals and dams. Amitrole is currently still registered in Australia for use against water hyacinth, but is less commonly used.
Other treatments
In Australia, manual and mechanical methods of controlling water hyacinth have been less popular than chemical methods of control. First, they are even less likely to achieve eradication of the weed than is treatment with herbicides, and second because they may result in little more than a culling of the plant population which causes a return to the exponential growth phase of the weed infestation. The recent advent of large harvesting machines combined with the growing public concern over the use of agrochemicals can be expected to result in a resurgence of interest in mechanical control of water hyacinth.
Discharge of sewage effluent, waste from industries and urban areas and leaching of nutrients from fertilized land into water systems promote nutrient-rich conditions favouring rapid growth of water hyacinth. Similarly, construction of dams, canals, lakes and reservoirs for agricultural, domestic, industrial and recreational purposes further increase the potential for problems from several aquatic weeds including water hyacinth.
Biological control
Arthropods
In its native range, E. crassipes is attacked by a complex of arthropods (Perkins 1973). Most interest in biological control of water hyacinth is currently centred on just six arthropod species.
The South American weevils Neochetina eichhorniae Warner and N. bruchi Hustache feed on the lamina and petiole as adults and deposit eggs usually on or near the young central leaf. Larvae tunnel in the petioles and late instar larvae are frequently found in the rhizomes from which they exit to pupate in the roots.
Neochetina eichhorniae and N. bruchi were liberated in Australia in 1975 and 1990 respectively. Because N. eichhorniae and N. bruchi have been much studied over many years and their host-specificity is beyond question, they have been released in many tropical and sub-tropical countries (Julien 1992) and are currently central to a major collaborative biocontrol project against water hyacinth in south-east Asia initiated by the Australian Centre for International Agricultural Research (ACIAR).
Aquatic vertebrates
Water hyacinth can be eaten by the manatee, Trichechus spp. (Bengston 1983) and the grass carp or white amur, Ctenopharyngodon idella (Cuvier and Valenciennes) (Baker et al. 1974, Delfosse et al. 1976).
|
References
Australia’s Virtual Herbarium, < http://www.anbg.gov.au/avh.html>, Commonwealth State and Territory Governments of Australia and the Council of Heads of Australian Herbaria, accessed July 25th 2006.
Baker, GE, Sutton, DL and Blackburn, RD 1974, ‘Feeding habits of the white amur on water hyacinth’, Hyacinth Control Journal, vol. 12, pp. 58-62.
Barrett, SCH 1977, ‘Trystily in Eichhornia crassipes (Mart.) Solms. (Water hyacinth)’, Biotropica, vol. 9, pp. 230-238.
Barrett, SCH and Forno, IW 1982, ‘Style morph distribution in New World populations of Eichhornia crassipes (Mart.) Solms-Laubach (water hyacinth)’, Aquatic Botany, vol. 13, pp. 299-306.
Bengston, JL 1983, ‘Estimating food consumption of free-ranging manatees in Florida’, Journal of Wildlife Management, vol. 47, pp. 1186-92.
Das, RR 1969, ‘A study of reproduction in Eichhornia crassipes (Mart.) Solms’, Tropical Ecology vol. 10, pp. 195-198.
Delfosse, ES, Sutton, DL and Perkins, BD 1976, ‘Combination of the mottled water hyacinth weevil and the white amur for biological control of water hyacinth’, Journal of Aquatic Plant Management, vol. 14, pp. 64-67.
Julien, MH 1992, Biological control of weeds: a world catalogue of agents and their target weeds, 3rd edn, CAB International/ACIAR.
Manson, JG and Manson, BE 1958, ‘Water hyacinth reproduces by seed in New Zealand’, New Zealand Journal of Agriculture, vol. 96, p. 191.
Matthews, LJ, Manson, BE and Coffey, BT (1977, ‘Longevity of water hyacinth (Eichhornia crassipes (Mart.) Solms.) seed in New Zealand’, Proceedings 6th Asian-Pacific Weed Science Society Conference1968, Volume 1, pp. 263-7.
Obeid, M and Tag el Seed, M 1976, ‘Factors affecting dormancy and germination of seeds of Eichhornia crassipes (Mart.) Solms. from the Nile’, Weed Research, vol. 16, pp. 71-80.
Penfound, WT and Earle, TT 1948, ‘The biology of the water hyacinth’, Ecological Monographs, vol. 18, pp. 447-472.
Perkins, BD 1973, ‘Potential for water hyacinth management with biological agents’, Proceedings Tall Timbers Conference, Ecological Animal Control Habitat Management 1972, pp. 63-74.
Pieterse, AH 1978, ‘The water hyacinth (Eichhornia crassipes) – a review’, Abstracts. Tropical Agriculture 4(2), 9-42.
Ueki, K and Oki, Y 1979, ‘Seed production and germination of Eichhornia crassipes in Japan’, Proceedings 7th Asian-Pacific Weed Science Society Conference 1979, pp. 257-260.
Ultsch, GR 1973,. ‘The effects of water hyacinth (Eichhornia crassipes) on the microenvironment of aquatic communities’, Archiv fuer Hydrbiologie, vol. 72, pp. 460-73.
Wright, AD and Purcell, MF 1995, ‘Eichhornia crassipes (Mart.) Solms-Laubach’, in Groves, RH and Shepherd, RCH and Richardson, RG (eds.) The Biology of Australian Weeds, Volume 1. RG and FJ Richardson, Frankston, Victoria, pp. 111-121.
Jatropha gossypiifolia L.
Name
Jatropha gossypiifolia L. (bellyache bush) is a member of the Euphorbiaceae family of plants, a very large family of world-wide distribution, containing many useful, weedy and poisonous plants among its 300 genera and about 7500 species. Its name is a combination of the Latin gossypium, cotton, and folium, leaf, suggesting that the leaves appear similar to those of the cotton plant. ‘Bellyache bush’ refers to the medicinal and purgative properties of the plant.

Figure 18.7a Bellyache bush, Jatropha gossypiifolia

Figure 18.7b an erect shrub, bellyache bush grows to a height of 2 to 3 metres
Description
Bellyache bush is an erect shrub or small tree with one to several stems rising from the herbaceous crown, 2 to 3 m high and reproducing by seed and suckers from the root.
There are one to several thick, rather soft stems, which are 1 to 2 m long and coarsely hairy. The stems exude a watery sap when injured.
The plant’s leaves are deep purple and sticky when young, bright green when mature. They are alternate, stalked, rounded or obviate, about 10 cm in diameter and deeply divided into 3 lanceolate lobes.
The margins and stalks are covered with small purple flowers with yellow centres, in clusters on branched stalks in upper leaf axils.
The fruit of bellyache bush is an oblong, 3-lobed capsule about 1.2 cm long and 1 cm wide. Seeds are dark coloured, about 8 mm long and the root is rather fleshy and tuberous.

Figure 18.8 Seedling and flowers of bellyache bush
History
A native of the Caribbean region, bellyache bush has been introduced as an ornamental to a number of countries including Africa, Madagascar, India, Indonesia and Papua New Guinea. It was introduced to northern Australia as an ornamental in the late 1800s. Over the intervening years it has become more aggressive and now occurs as a relict weed around abandoned homesteads and mine shafts and is encroaching onto degraded rangelands. It was listed as naturalised in Queensland in 1912. On Queensland’s list of priority weeds, bellyache bush is ranked number three, behind rubber vine and parthenium.
Distribution
Bellyache bush is naturalised at Charters Towers, Townsville and other places in Queensland and also occurs at Darwin, Middle Point, Batchelor, Katherine, Willeroo, Mataranka, Daly Waters and the Barrys Caves on the Barkly Highway in the Northern Territory. It is also scattered along the Great Northern Highway between Wyndham, Halls Creek, Fitzroy Crossing and Derby, continuing south along the coast from Cape Leveque to Cape Bosset in tropical Western Australia.
Its distribution is in tropical savannah areas with distinct wet/dry seasons. It does not occur in the southern half of Australia.

Figure 18.9 The Australian distribution of bellyache bush (Australian Virtual Herbarium, 2006)
Habitat
Bellyache bush is well adapted to areas that have a pronounced dry season, and its distribution has expanded despite dry conditions over the past few years. The plant is very drought tolerant. Its potential range is the entire tropical savannah.
Bellyache bush invades disturbed habitats – particularly rangelands and riparian zones. Its growth is much faster out in the open than under the canopy.
Growth and development
Seeds germinate with the break of season between October and December. Seedling plants develop at a moderate rate during the remainder of the wet season, slowing in the following dry winter unless there is ample groundwater close to the surface. The age at first flowering is not known with certainty in Australia but seems to be at least two years. Once flowering commences, however, it continues throughout the year. Established plants form new shoots from the crown and roots at the break of season.

Figure 18.10 Flowers of bellyache bush
Reproduction
In a non-stressed situation, bellyache bush may flower 55 days after germination. Increased competition/moisture stress leads to increased time to flowering (up to more than a year). Male and female flowers occur on the same plant, and insects are essential for normal seed set.
Bellyache bush can regenerate from stem cuttings – it is semi-succulent and deciduous. Stem cuttings can produce viable seed from the stored energy in the stem. The plants shed leaves in the dry season due to moisture stress or cold.
Plants can grow up to 2 metres annually, but more commonly grow 0.5 to 1 metre annually. In native areas plants seldom reach more than 30 cms high.
Fruiting is largely rainfall dependent. It begins in September/October and may last up to 10 months. The period from flowering to seed maturity ranges from 18-32 days, depending on competition.

Figure 18.11 Leaves of young bellyache plants have a distinctive deep purple colour
The seed capsules explode when mature, throwing the seed up to 13 metres. Floods can move plant material and it readily takes root and establishes. Seed dispersal can be facilitated by meat ants over short distances and by humans over long distances. Seed bank survival can be 4 years, and in Queensland the plants can survive up to 15 years.
With the increasing quantity of seed becoming available, bellyache bush is tending to form dense thickets, crowding out useful species and reducing productivity in some Northern Territory areas. All parts of bellyache bush are highly toxic and can kill grazing animals if ingested. The seeds are also toxic to humans.
Weed management
Herbicides
There are two main options for chemical management of bellyache bush, Fluroxypur and Metsulfuron. Fluroxypur effectiveness decreases with seasonal leaf drop, but Metsulfuron is effective all year round. An aerial application trial at Home Hill and Palmer River (most northern of Queensland infestations) was effective, but non-target species such as Melaleuca were impacted. Follow up control is important as the recruitment is good.

Figure 18.12 a field site dominated by bellyache bush
Other treatments
Competition – well pastured paddocks will contribute towards increased bellyache bush mortality, reduce growth and delay the onset of reproduction for up to 4 years.
Cut stump – ground level cutting/slashing is effective year round.
Burning – a 10 second burn around the base of the plant will control bellyache bush plants.
Flooding – inundation with water over 1 – 2 weeks will kill bellyache bush plants.
Annual follow up will need to occur for 4 – 6 years to achieve seed bank depletion.
Biological control
A moderately large suite of natural enemies attack bellyache bush in its native range. There have been 47 species identified as potential biocontrol agents, all of them insects except for a rust fungus. Agonosoma trilineatum (the bellyache bush jewel bug), which sucks the sap of seeds, was imported in 2000. Specificity studies have been completed and it was first released on bellyache bush in 2003, but hasn’t yet established.

Figures 18.13a and b: Adult jewel bug (left) and nymphs, potential biological control agents for bellyache bush
Rejected biocontrol agents include:
Largocheirus spp – because they feed on cassava (a potential crop plant)
Colaspis sp – difficult to rear
Cylindrocopturus – difficult to rear
Rust fungus (Phakopsora jatrophicola), which attacks the leaves of bellyache bush, is being reared in the UK due to a high risk of escape from quarantine. Further funding is required to continue investigation into suitability.
Agent release in Queensland
Over 24000 robust adults of Agonosoma trilineatum were released in 2003 at sites close to Charters Towers, along the Burdekin River and along the Palmer and Walsh Rivers. Nymph survival is variable – on average 30 %, but can be as high as 90 %. A small number have been recorded at some sites. Reduced available seed over winter could be the cause of low survival.
Agent release in NT
There have been 75 releases of Agonosoma trilineatum. No establishment was observed. Release in cages was attempted but proved difficult because the plants need pollinators to produce seed.
Summary
Bellyache bush is a species for which a lot is known about its biology, ecology and field management techniques, but surprisingly little about both ecological and economic impacts. Better data on the rate of spread is needed to assess the impact of biocontrol, and to infer containment strategies. Climex modelling has shown promise in narrowing down where surveys should be undertaken in what is a very broad native range – consensus appears to be that South America is best.
References
Dhileepan, K and Harvey, S (editors) 2007. Workshop Proceedings – Bellyache Bush Research Workshop. Alan Fletcher Research Station, DPI&F, Sherwood, Qld.
Parsons, WT and Cuthbertson, ET 1992 Noxious Weeds of Australia. Inkata Press, Melbourne.
Useful websites:
www.csiro.au/science/BellyacheBushBiocontrol.html
Anredera cordifolia L.
Name
Anredera cordifolia (madeira vine) is an aggressive perennial creeper belonging to the Basellaceae family. The plant is also known as Potato Vine and Lamb’s Tails. It is a native of South America and was imported because of its attractive white flowers and strong growth. It was often planted beside outdoor latrines in Australia, as the leaves, when eaten, were believed to have a laxative effect.

Figure 18.14 Madeira vine
Madeira vine is an aggressive, rampant twining perennial climber from South America, which races to the canopy, curtaining all vegetation with its thick rope-like stems and dense foliage.
It has fleshy, heart-shaped hairless leaves, tips rounded or shallowly indented, base lobed. Leaves are light green underneath and darker on top. Stalks are often reddish.
The vine produces masses of perfumed creamy white tubular flowers between March and May. The small fragrant cream flowers are arranged in slender racemes (lamb’s tails), which droop from leaf axils. The flowers are short-lived and fruit rarely forms. A curtain of leaves and flowers is often formed.

Figure 18.15 Flowers of Madeira vine
Stems bear thousands of long-lived aerial tubers which form clusters high in the vine. Underground tubers, which may be football sized, grow on rhizomes up to a metre deep.
History
Madeira vine is native to South America and was probably imported because of its attractive white flowers and strong growth. The plant came into Australia about 100 years ago and was naturalised in Queensland about 55 years ago.
Originally a garden escapee in NSW and Queensland, which has been suspected of poisoning stock, it has also been recorded on the north-western slopes of NSW and is now present throughout Australia. Under climate change Victoria could also become a more serious location for the vine.
Madeira vine is a problem in Australia, New Zealand, Hawaii and South Africa.
It is the fifth worst environmental weed in Queensland; 41 out of 71 in WONS. Madeira vine is a declared noxious weed in some council areas in NSW, and is a declared Class 3 plant under the Land Protection (Pest and Stock Route Management) Act 2002.
Distribution
Madeira vine is now widespread. Adapting to a vast range of soils and conditions, Madeira vine has spread along waterways and through forests and is now considered to be the greatest weed threat to rainforest remnants.
It is a serious weed that will invade the margins of rainforest smothering small trees and shrubs. It is evidently partly salt tolerant, as it has been noted overgrowing mangroves.
The vine is also spreading along inland watercourses in NSW and along the north-western slopes of NSW.

Figure 18.16 Current and potential distribution of Madeira vine in Australia
Habitat
Madeira vine prefers warm, damp or seasonally moist locations with lightly shaded or sunny conditions. It has been used domestically for screening fences, sheds etc.
It invades the edges of bushland, woodland and rainforest. Infestations rapidly expand as existing vegetation is smothered.
Tolerant of drought, salt and frost, it will die back to the tuber and reshoot.
It grows from sea level up to 3000 m, but is best where there is 500 – 2000 mm average annual rainfall.

Figure 18.17 Rainforest remnant covered by Madeira and other exotic species
Growth and development
Madeira vine is very fast growing and can cover other plants quickly. It produces numerous stems, reaching from the ground up into the canopy. Mature stems are woody with wart-like growths and can reach up to 20 metres.
Madeira vine can grow up to one metre of stem within one week during the summer and the tubers can be viable for up to 10 years. Tubers are produced on the stem at the base of the leaf and vary in size from ‘pea’ size to large clumps of tubers resembling ginger and can be ‘sweet potato’ size underground.

Figure 18.18 Stem tubers
Reproduction
Madeira vine reproduces vegetatively, via prolific production and subsequent dispersal of multitudes of subterranean and aerial tubers. These tubers are dispersed primarily by humans deliberately dumping waste and also by water, wind, gravity and soil disturbance from rodents, bush turkeys etc.
The vine grows easily from both kinds of tubers and also from pieces of rhizome. The tubers are very easily dislodged from the parent vine, and both tubers and rhizomes can be washed down waterways.
The tubers can remain viable for 5 to 10 years, with anecdotal reports of persistence for up to 15 years.
Seed and fruit production are considered rare in our climate, but fruit production has been reported in the native range – fruit bearing populations have been identified in Toowoomba and the Lockyer Valley.
Importance
Madeira vine is a devastating weed which invades moist forest and rainforest edges. It blankets the ground and envelops the canopy. The vine restricts light, encourages disease and prevents germination of native plants. Its weight can break down trees and Madeira vine also helps to destroy rainforest.
Weed management
Physical/mechanical control
This type of control is only practical for small infestations. All vines must be severed and removed. Prior to severing vines a tarpaulin or plastic sheet should be placed below to catch aerial tubers and as many aerial tubers as possible should be removed. Subterranean tubers and roots should be dug up and follow-up is required within 3 months.
Tubers and all plant material should be disposed of by:
- incinerating in a hot fire
- placing in plastic and leaving in the sun to ‘cook’ (leave to compost for two years)
- decomposing in a covered drum of water
- freezing
Herbicides
Madeira vine is very hard to kill with herbicides because of its tubers – ideally remove every tuber from the trees and ground first.
Spraying – Herbicide treated plants tend to die back to the tuber, with resprouting occurring from the next tuber section. Follow-up spraying is essential and should be initiated when regrowth from the tuber has sufficient leaf area to assist in the translocation of herbicides (if the leaves are allowed to re-grow for too long, new tubers will be formed underground).
Metsulfuron-methyl + wetting agent is a selective herbicide and affects plants differently. Foliar spraying may not be the best method if native plants are affected by the herbicide, but is useful where the weeds have formed an impenetrable mass as the dense weed canopy protects any submerged natives.
Scrape and paint method – Scrape 30 cm sections along the vine stem. As you scrape, immediately paint each section with 100 % glyphosate or picloram gel. Treated aerial tubers and vines are left in the canopy to decompose.
Biological control
In Queensland Madeira vine was ranked fifth best weed to target for BC – it is the only plant in Basellaceae in Australia, but little is known about the plant. Several agents have been tested in Argentina and one in South Africa. One agent, Plechtonycha correntina (a leaf feeding beetle) is already at AFRS – both larvae and adults feed on the plants. One Asian vegetable Basella alba (Ceylon spinach) may be a problem – this is a climbing perennial herb grown as a leaf vegetable rich in nutrients and vitamins. It is grown on a small scale by the vegetable industry and may be vulnerable to Plechtonycha correntina.
Summary
Madeira vine is a devastating weed capable of smothering and obliterating all indigenous vegetation. Control of this plant is difficult and success requires diligence and patience as well as many follow-up visits and repeated treatments. The impacts of climate change on Madeira vine distribution, as well as population dynamics and rate of spread without intervention need further research. An integrated approach to Madeira vine management is needed, especially if an effective biocontrol is found.
References
Auld, BA and Medd, RW 1987 Weeds: an illustrated botanical guide to the weeds of Australia. Inkata Press, Melbourne.
Taylor, D and Harvey, S (editors) 2007 Workshop Proceedings – Madeira Vine Research Workshop. Alan Fletcher Research Station, DPI&F, Sherwood, Qld.
Useful website:
Mikania micrantha H.B.K
Name
Mikania vine (Mikania micrantha) is a member of the Asteraceae family. It is also known as ‘wa butako’ in Fijian (vine that steals or invader) and ‘green tidal wave’ in India for its smothering habit on tea plantations.

Figure 18.19 Mikania micrantha
Description
Mikania vine is a rapidly growing perennial creeper. It is an herbaceous, sprawling twining vine that smothers other vegetation and infrastructure.
Mikania vine leaves are smooth and heart-shaped. They are 4-13 cm long and are arranged in opposite pairs along the stem. There are three main veins that arise from the base of the leaf. The stems are slender, ribbed and bear fine, white hairs – lateral stems are as vigorous as the main stem.
The flowers of Mikania vine are white to greenish-white and are produced in clusters mainly at the ends of stems growing in full sunlight. Each flower head is 4-6 mm long and contains four individual flowers.
History
Mikania vine is native to Central and South America. It has become a serious weed in West Africa through to India, South-East Asia, Indonesia and also the Pacific Islands.
Mikania vine was first found in Australia in 1998 at Ingham and Bingil Bay.
Distribution
Mikania vine is native to Central and South America and has become a serious weed in West Africa through to India, South-East Asia, Indonesia and also the Pacific Islands.
Populations have been detected in three shires in Queensland: Hinchinbrook Shire (Ingham), Johnstone Shire (Bingil Bay and Mission Beach) and Mareeba Shire (Speewah).
The potential distribution in Australia includes the coastal regions of the Northern Territory and much of eastern Queensland, extending into north-eastern New South Wales.

Figure 18.20 Native and introduced range of Mikania micrantha
Habitat
Mikania vine thrives in humid areas where rainfall exceeds 1000 mm per annum and prefers areas with rich, damp soils. It grows well in open areas as well as in partial shade and disturbed areas.
The vine will also grow in deep shade although it is unlikely to flower in these areas. It can, however, grow up through a closed forest to the canopy.
Growth and development
Mikania vine is a rapidly growing, perennial vine with many branches. From a single plant, it can smother an area of 25 square metres in months.
The vine flowers from May to June and from September to October, but can also flower in between these periods (July and August). Plants will also flower heavily out of season in areas exposed to full sunlight. Each flower head is 4-6 mm long and contains four individual flowers.

Figure 18.21 Mikania vine will grow rapidly in the wet season and may die back to ground level in the dry season.
The lateral shoots will twine around the vine’s own main stem until other support is found. In the absence of support, Mikania vine is prostrate and will form roots along any stem that is touching the ground.
Reproduction
Mikania vine has the ability to reproduce both by seed and vegetatively. The vine produces tufted seeds which are equipped for wind dispersal. Each plant can produce 40,000 seeds per year. The seeds are black, 1.5-2 mm long, thin and five-angled. The seed tufts (pappus) consist of over 30 fine white hairs or bristles. The pappus is longer than the seed itself. The seed tufts can be transported by wind, water, machinery and animals.

Figure 18.22 Mikania seed and vegetative reproduction
Mikania vine is also very effective at vegetative reproduction. Small plant fragments which contain a node will produce roots when in contact with moist soil. Likewise, roots will form along a stem where it touches the ground.
Importance
If it becomes established in Australia, Mikania vine will damage the natural environment and affect the habitat of native animals. It has the ability to spread rapidly and smother native vegetation and crops. It would therefore pose a major threat to the Wet Tropics World Heritage areas in Queensland.
It can also cause serious damage to agricultural industries in the tropics, including sugar cane, tropical fruit and vegetable production.
When mixed with the soil, Mikania vine debris produces toxins that inhibit growth of vegetation, such as native plants and agricultural crops.

Figure 18.23 Mikania vine is a Class 1 Declared plant in Queensland – it is not commonly present, but if introduced would cause serious economic, environmental or social impact.
Weed management
Herbicides
Where appropriate, herbicides are used to reduce the size and density of Mikania vine infestations.
Table 18.1 Effect of herbicides used in various countries
| Herbicides | Country | Effect | Reference |
| 2,4-D | India | Effective | Palit, (1981) |
| Bentazone and Picloram | China | Effective | on inhibiting seed germination Hu and But, (1994) cited in Zhang et al.,(2004). |
| Imazapyr | Malaysia | Uptake of Imazapyr ↑ with ↑ in light intensity | Ipor and Price, (1992). |
| Paraquat | Malaysia | Uptake of paraquat↓ with ↑ in light intensity | Ipor and Price, (1994). |
| Diuron and Oxyfluorfen | India | Inhibit germination of seeds | Abraham, (2002). |
| 2,4-D amine Paraquat Mixtures combining 2, 4-D amine + Paraquat | Malaysia | Effective, used by farmers | Teoh et al.,(1985).Seth, (1971). |
| Grason DS (triclopyr + picloram) | India | Reduce biomass by 95 % | Sreenivasan and Sankaran (2001). |
| Glyphosate | India | Reduce biomass by 90% | Sreenivasan and Sankaran (2001). |
| Paraquat | Fiji | Effective, used by farmers | Robinson and Singh, (1973). |
Other treatments
The most effective way to control Mikania vine is to prevent its spread in the early stages of infestation. Small plants can be completely destroyed by manual removal after herbicide treatment, and by preventing contact with the soil. The roots of large plants can be removed and destroyed. It is important that all plant fragments must be removed and destroyed, as the vine will grow from plant fragments. Fire can be used to control large infestations of the vine.
Biological control
In 2007 14 infestations were identified and contained in Australia. There are several boological control agents which are in use overseas:
Actinote thalia pyrrha: this butterfly lays its eggs on the underside of Mikania vine leaves. Only the larvae feed on the leaves.

Figure 18.24 The Actinote butterfly larvae feed on Mikania leaves
Fungus: Puccinia spegazzinii de Toni. Young plant tissue is most susceptible to this rust fungus – often merges to form elongated pustules.

Figure 18.25a and b Young plant tissue is most susceptible to Puccinia
References
Biosecurity Queensland: Fact sheet PP143 – Invasive plants and animals – Mikania vine. Department of Primary Industries and Fisheries, Queensland, May 2007.